LA ANTÍTESIS NEODARWINISTA
Y OTROS ENSAYOS BREVES SOBRE EVOLUCIÓN
Daniel Heredia Doval
Universidad Autónoma de Madrid. E-mail: Esta dirección de correo electrónico está siendo protegida contra los robots de spam. Necesita tener JavaScript habilitado para poder verlo.
ÍNDICE
- Introducción
- La Antítesis Neodarwinista: Selección, Variación y Éxito
- Las fuentes de Variación: Mecanismos generadores de biodiversidad
- Revisando la Adaptación: Problemas actuales y una propuesta alternativa
- Evolución, Especiación y Adaptación: Visión personal general
- Agradecimientos y Bibliografía
Introducción
En este trabajo se compilan una serie de ensayos breves sobre la evolución de las especies, la insostenibilidad de su explicación ortodoxa y la necesidad de crear un modelo complejo capaz de explicar, en la medida de lo posible, la gran biodiversidad que observamos de forma empírica y coherente.
Aunque mi intención inicial era la de exponer un mayor número de ensayos y desarrollar en mayor profundidad las cuestiones que finalmente constituyen este trabajo (por razones de tiempo y organización ésta es la extensión final), espero haber podido plasmar eficientemente los aspectos fundamentales de mi pensamiento.
En todo caso, el objetivo de este trabajo no ha sido otro que el de poder pensar, organizar, y redactar mis ideas actuales sobre la evolución y el origen de la diversidad de las especies.
Puesto que los ensayos fueron escritos de forma discontinua, no existe un orden ni consecuencia entre ellos, sino que pretenden abordar distintas cuestiones sobre el mismo tema, aunque, en ocasiones su extensión se solapa.
Finalmente, he de expresar que considero la integración multidisciplinar e ideológica en la evolución la manera de hacer frente a los nuevos desafíos de la biología para tratar de comprender así, en la medida de lo posible, las enormes dimensiones de la vida: un fenómeno que tiende a diversificarse y autoconservarse, creando redes cada vez más complejas entre sus componentes. La evolución de las especies, así como el sexo o la transferencia horizontal sirven a este fin, la perpetuación de este fenómeno emergente que somos nosotros, las miles de bacterias de nuestro intestino y la totalidad de nuestro planeta simbiótico.
Daniel Heredia Doval.
Agradecimientos
Quiero agradecer su colaboración en la elaboración de las ideas de este trabajo, así como a su síntesis a Dunia Aguirre y a Arturo Sastre. Gracias a ambos por aquellas charlas en cualquier situación.
Bibliografía
Libros:
-Deconstruyendo a Darwin. Javier Sanpedro.2002
-Dientes de Gallina y dedos de caballo. Stephen J. Gould
-El planeta simbiótico. Lynn Margulis
-Evolución. Origen, adaptación y divergencia. Fontdevila y Moya
-Genes VII. Lewis
-La tautología darwinista. Fernando Vallejo
-Lamark´s Signature. Steele, Lindley y Blanden.
-Lamark y los mensajeros. Máximo Sandín.
-La sonrisa del flamenco. Stephen J. Gould.
-Tratado de botánica. Strasburger.
-El origen de las especies, Charles Darwin.
Artículos:
-Large-Scale Variation Among Human and Great Ape Genomes Determined by Array Comparative Genomic Hybridization, Devin P. Locke et al, 2003.
-Mobile Elements: Drivers of Genome Evolution, Haig H. Kazazian Jr. 2004.
-Bonemaking Protein Shapes Beaks of Darwin’s Finches, Elisabeth Pennisi,2004.
-The future of selection: individuality, the twin legacies of Lamarck & Darwin, Hugo Hoenigsberg, 2002
-Brazilian Journal of Genetics. Vol. 20 nº4
-Current Opinion in Genetics & Development. Genomes and evolution.
-Initial sequencing and analysis of the human genome, International human sequencing consortium, 2001
-Patchwork people, Erika Check 2005
-La transformación de la evolución, Máximo Sandín, 2004
On-line:
-Revista de evolución del GEB (UAM). www.uam.es/otros/geb/revista/Portada.htm
-Enciclopedia multimedia encarta 2002.
La Antítesis Neodarwinista
Sobre la Variación, la Selección y el Éxito
El Neodarwinismo es la teoría evolutiva más aceptada en la actualidad. Su origen se remonta a principios del siglo XX y es el resultado de la fusión de los conceptos de selección de la teoría darwinista y los conocimientos de genética de poblaciones de la época, basados en experimentos matemáticos aleatorios sobre la sustitución de alelos.
Esta teoría está fundamentada en tres pilares básicos: Variación, Selección y Éxito. Según la doctrina neodarwinista, las mutaciones puntuales y al azar generan pequeñas diferencias entre los individuos de una especie (Variación), de forma que, en un estado de constante competición por los recursos, sólo sobreviven los individuos más adecuados (Selección Natural) y/o son estos los que dejan más descendencia (Éxito reproductivo) de forma que sus descendientes y las características que aportan esa mayor eficacia acaban siendo las normales de la población. Este supuesto prolongado durante un amplio espacio de tiempo y aplicado a innumerables caracteres daría lugar a la formación lenta y gradual de especies nuevas y a una adaptación continua de las mismas.
Ahora bien, ¿pueden realmente estos pilares sostener a la teoría sintética? es más, ¿son sostenibles todos ellos en si mismos en la naturaleza?
Sobre la Selección Natural
La Selección Natural es la verdadera aportación de Darwin a la teoría evolutiva hegemónica. La Selección Natural es la fuerza que decide quien vive y quien muere, es la presión a la que se ven sometidos los individuos como resultado de las dificultades del medio y de las relaciones con otros seres vivos, eliminando a los menos afortunados y recompensando a los más adecuados. ¿De dónde surgió este precepto sospechosamente empresarial?
Charles Robert Darwin (1809-1882) nació en Shrewsbury (Inglaterra), el 12 de febrero de 1809. Fue el quinto hijo de una acomodada y sofisticada familia inglesa. Su abuelo materno fue el próspero empresario de porcelanas Josiah Wedgwood y su abuelo paterno fue un exitoso médico, Erasmus Darwin.
Tras terminar sus estudios de Teología, habiendo abandonado la carrera de medicina anteriormente, se enroló en el barco de reconocimiento HMS Beagle, con el cual visitó, entre otros lugares, las famosas islas Galápagos, tomando apuntes y recogiendo muestras. Cuando volvió a Inglaterra no tenía una idea clara sobre el origen de la diversidad de las especies en las distintas islas que visitó. Finalmente, tal como consta en “El Origen de las Especies”, usando los conceptos de Thomas Robert Malthus (Ensayo sobre el principio de la población, 1798) que expone que en las poblaciones humanas nacen más individuos de los que pueden sobrevivir; influido por la corriente geológica del gradualismo (de Charles Lyell) y apoyándose en los conocimientos que tenía sobre ganadería y agricultura, así como de la selección antrópica a la que se someten a las especies criadas para el consumo humano (reses, cultivos y animales de compañía), Darwin desarrolló su teoría y propuso el término de Selección Natural como un mecanismo de cambio y regulación de las poblaciones.
Tras esta reflexión histórica centrémonos en la validez de este omnipotente concepto: ¿puede realmente una extrapolación de la actividad humana explicar toda la complejidad de la naturaleza? Aun siendo obvio el hecho de que unos individuos viven y otros mueren, ¿este hecho surge realmente de la acción de una indetectable fuerza selectiva?
Descartando a los seres incompatibles con la vida, aquellos que sufren graves afecciones y mueren, todos los individuos de una población son en si “adecuados”. Las poblaciones tienden a un morfotipo normal perfectamente adaptado a su ambiente, de forma que este ha de ser difícilmente superable por una aberración. Pero es más, las posibilidades de supervivencia, así como de reproducción, no dependen de forma exclusiva del genotipo, que es en esencia similar, sino que recaen en las interacciones con el ambiente y las circunstancias de su existencia; de modo que, por apto que sea, un individuo puede ser devorado antes de nacer, romperse una pata, sufrir una hambruna, caerle un rayo…¿es esto también selección natural?
A mi entender solo hay dos tipos de individuos en las poblaciones naturales: no aptos para la vida y potencialmente aptos para la vida. No hay gradaciones.
Sobre la Competencia y el Éxito
La supervivencia y el mayor éxito reproductivo son las recompensas que aguardan a los más adecuados en un estado de constante competición inter e intraespecífica. Sin embargo, ¿no pueden ser estos supuestos hechos burdas interpretaciones humanas del mundo natural?
La competencia feroz, motor del neodarwinismo, no es el estado normal de las poblaciones naturales, que en esencia tratan de vivir bajo la compleja red de interacciones naturales sin preocuparse por la aptitud de sus vecinos. En las poblaciones naturales, no nacen más individuos de lo necesario, puesto que el número de crías está determinado por la interacción con el ambiente, adecuado a las dificultades que pueden tener los juveniles en llegar a adultos y regulado por la dinámica del ecosistema (predadores, alimento, otros recursos…). La supervivencia de los individuos depende por lo tanto más de su estado de salud y edad que de ningún otro factor. La competencia feroz no es sino el subproducto de una alteración en la dinámica normal de un ecosistema. Así pues, solo cuando existe una superpoblación y/o una escasez de recursos (de forma abrupta) los individuos se pueden ver forzados a competir de forma evidente por su supervivencia, y aun en este supuesto, la selección natural no tiene mayor poder de explicación que el mero azar.
Por otra parte está la selección sexual, que es un concepto derivado, como muchos otros, de la selección natural, dicta que en especies con reproducción sexual, un individuo de un sexo (normalmente la hembra en los animales) elige a su pareja en función de su aptitud, y en consecuencia, los individuos más adecuados se reproducen con más éxito y dejan mayor descendencia. Sin embargo, a mi entender, cuando un individuo elige a una pareja tras un ritual o bien por su aspecto, no esta midiendo su aptitud en sentido neodarwinista, sino que mide su nivel de salud y bienestar físico. Seleccionando una pareja tras un ritual, una hembra se asegura que no perderá el tiempo (como mínimo) apareándose con individuo mal nutrido, enfermo o incapaz de suplir sus necesidades. La diferencia de elegir por aptitud o por sanidad radica en el hecho de que la salud varía con el tiempo, y un individuo puede no estar preparado para reproducirse un año, pero probablemente se reproducirá a lo largo de su vida si mejoran sus condiciones sanitarias o alimenticias, y no existe por lo tanto una evidente y justificada reproducción diferencial entre los individuos
Sobre la mutación, variación alélica y la especiación
Desde el punto de vista del Neodarwinismo ortodoxo, solo hay una fuente posible de variación: la mutación puntual al azar. Una mutación puntual es un cambio discreto en una secuencia génica que puede afectar, o no, a la proteína final. Si el efecto no es neutro, la proteína suele quedar dañada y puede afectar al individuo en mayor o menor medida. Las macromutaciones (entiéndase cualquier cambio genético superior a unos pocos pares de bases), difícilmente pueden dar lugar a un pequeño cambio fenotípico, por lo que no pueden ser consideradas como una fuente de cambio gradual, y un individuo con grandes diferencias frente al resto de individuos normales no será capaz de reproducirse y no podrá pasar su alteración a la descendencia.
Para la visión neodarwinista, una mutación puede ser neutra, perniciosa o beneficiosa para el individuo y crea una variedad donde seleccionar. De esta forma, los individuos con mutaciones neutras no sufren selección alguna, puesto que el cambio en la secuencia del gen no afecta a la proteína que produce, mientras que los individuos con una mutación perniciosa o incompatible con la vida son “seleccionados negativamente” (es decir, mueren) y los individuos con una mutación beneficiosa para su supervivencia son “seleccionados positivamente” (sobreviven y se reproducen más). En este último supuesto, la mutación se expande por la población, pasando a ser un alelo nuevo (variante del gen) que acabará sustituyendo al original, con el tiempo.
Sin embargo, las mutaciones al azar son realmente raras en las poblaciones naturales y los seres vivos poseen diversos mecanismos de reparación, comprobación y taponamiento para evitar su efecto. Las mutaciones que afectan a la proteína final suelen invalidar este producto de forma que o bien impiden el desarrollo correcto de su función directa o bien provocan efectos en las interacciones génicas. Normalmente el resultado visible (cuando es no letal) es una afección hereditaria (ej: hemofilia) o una anomalía de poca gravedad (ej: albinismo). Sólo se conoce un caso en el cual una mutación es “beneficiosa”: en las poblaciones africanas, los individuos heterocigotos para la anemia falciforme parecen tener mayor resistencia a la malaria. Sin embargo, los homocigotos mueren de anemia, por lo que igualmente la mutación no se expande de forma efectiva por la población.
Por otra parte, en una población de gran tamaño la sustitución gradual de alelos de forma “natural” es en si imposible. Suponiendo que una nueva variedad alélica que tendiese a difundirse por la población cumpliese todos los requisitos neodarwinistas (provoca una pequeña diferencia frente al fenotipo normal, genera una ventaja competitiva y no desaparece por causas externas), esta variedad alélica se cruzara infinidad de veces con el alelo salvaje, de manera que o es un carácter mendeliano de dominancia completa (lo cual es extremadamente raro) o, admitiendo la acción omnipotente de la selección natural, la ventaja que proporciona debería ser clara, o lo que es lo mismo, una innovación plausible y no gradual. Además, puesto que analizamos una población grande, el tiempo de sustitución de las variedades alélicas debería de ser muy prolongado.
La sustitución de alelos de forma eficiente parece no poder ser un proceso gradual, sino el resultado de un sesgo en la población donde, por diversas razones, una variedad pasa a ser la más representativa de la población (como en el efecto fundadores).
Por otra parte, hasta hace poco tiempo se pensaba que las variantes alélicas eran la única fuente de variabilidad dentro de una población, sin embargo, trabajos recientes e independientes ponen de manifiesto que gran parte (si no la mayoría) de las diferencias entre individuos no responden a diferencias alélicas de algunos genes sino al número variable de repeticiones que presentan de estos, principalmente relacionados con la respuesta al ambiente. Esto crea un problema logístico, puesto que empíricamente queda demostrado que las mutaciones puntuales al azar no son las principales generadoras de variedad, sino las duplicaciones seriales, que son difícilmente explicables por fallos de replicación y requieren de otros mecanismos implicados.
La especiación es la aparición de nuevas especies. Los retrocruzamientos continuos, la similitud estructural de los alelos y la limitación de tiempo son impedimentos cruciales para admitir la acumulación lenta y gradual de cambios puedan llegar hasta crear la incompatibilidad reproductiva entre dos morfotipos. Como resultado, un nuevo alelo no pernicioso capaz de expandirse por una población podría dar lugar a polimorfismos, es decir, variedades de un carácter, pero no a especies nuevas.
La especiación requiere de una incompatibilidad reproductiva, y esto sólo es posible cuando ocurre una alteración brusca en una serie de individuos que les impide reproducirse con los individuos “normales”. Las duplicaciones, la poliploidización, los reordenamientos cromosómicos y la incompatibilidad citoplasmática son, entre otros, cambios en los organismos capaces de crear aislamiento reproductivo. Son bien conocidos casos de especiación producidos por estos sucesos (sobre todo en insectos y plantas), mientras que (al menos que a mi me conste) no se han descrito casos de especiación por sustitución gradual de alelos.
Conclusión
Tras analizar por separado, de forma crítica, los pilares sobre los que se sustenta la teoría neodarwinista, a mi entender, parece claro que no es que haya que adaptar esta teoría a los descubrimientos recientes (que cada vez se ajustan menos al modelo “ideal”) o hacer pequeños ajustes, sino que la Síntesis neodarwinista tiene profundos problemas de base que no se pueden arreglar modificándola ligeramente, porque estos problemas derivan, desde su origen, de una mala interpretación de la naturaleza, del desconocimiento de la dinámica interna de los organismos, de la aplicación de ideales humanos un fenómeno natural y la construcción de un modelo inaceptablemente simple para explicar toda la complejidad de la vida.
Con la cantidad de información que poseemos, los apasionantes y sorprendentes nuevos descubrimientos sobre la actividad de los genomas (donde reside la información potencial de los individuos) y usando el sentido común, debemos de tratar de crear un nuevo modelo para tratar de explicar de forma coherente la biodiversidad en la naturaleza, sin caer en los errores que se cometieron en el pasado.
Problemas actuales y una propuesta alternativa
La adaptación es el proceso de adecuación al medio por parte de los organismos y sus resultados son de una exquisita precisión.
Desde el punto de vista de la teoría sintética, la adaptación es un proceso gradual resultante ¡como no! de la acción de la selección natural. Aquellos individuos mejor adaptados al ambiente se reproducen con mayor éxito y transmiten sus “ventajas” a la descendencia que, con el tiempo, se harán de los caracteres mayoritarios de la población para, finalmente, originar por acumulación de variaciones una nueva especie perfectamente adecuada a su entorno.
Este hecho implica que todas y cada una de las características de todas la especies, la increíble precisión con la que se coordinan y actúan todas sus partes, es producto de la acumulación de golpes de azar que, por supuesto, dieron por separado una ventaja tal que sustituyó al morfotipo original de la especie.
Solo de forma intuitiva resulta evidente que son demasiados caracteres, demasiada complejidad, demasiado azar y demasiado poco tiempo para agregar y fijar novedades. Sin embargo, pese a lo que indica la lógica, toda la biología evolutiva tiende a explicarse mediante este proceso, hasta el punto de llegar a “meter con una cuña” infinidad de casos de difícil explicación por esta idea. Los ejemplos simples de libro de texto pueden pasar por este modelo (las liebres blancas en la nieve y la polilla del abedul), otra cosa es como aparecen estos caracteres y como se fijan en realidad. Pero al tratar de explicar toda la biodiversidad por este supuesto fenómeno nos daremos de bruces con ejemplos incompatibles con nuestra propia lógica.
Este es el caso de nuestra especie, en el cual los antropólogos se empeñan en buscar ventajas adaptativas entre los distintos homínidos y e incluso en las distintas etnias. Tratar de explicar las diversidad humana y la adaptación de las distintas etnias al ambiente al que viven, bajo este modelo es completamente absurdo, puesto que en un grupo organizado de cazadores-recolectores (como se admite que es nuestra estructura social original) la supervivencia no es un aspecto individual, sino que depende de la interactuación y cooperación de todo el grupo. Por lo tanto, pese a cualquier variación individual, por mucha ventaja que proporcionase a la persona que la poseyese, esta tendría que cooperar y compartir los víveres que consiga con el resto del grupo, ayudando a la supervivencia de los “menos aptos” y a su vez dependiendo de estos para su propia supervivencia. Bajo este supuesto, ¿es posible la expansión de nuevas variantes alélicas hasta sustituir a la forma original? teniendo en cuenta que la diversidad humana actual se ha producido en escasos miles de años, parece “poco probable”.
Como ya se dijo, la visión ortodoxa trata de explicar la perfecta adecuación al medio de cada especie/población/ser por medio de un reducido trinomio de Azar + Selección + Tiempo, donde el último término depende directamente de la magnitud del problema a explicar. Resulta poco convincente en este y otros muchos ejemplos que este sea (al menos el único) mecanismo de adaptación.
¿No es mucho más intuitivo y lógico pensar que los organismos responden al ambiente implicando a su genoma (como es sabido por otra parte) y que esta respuesta puede fijarse en población, desterrando al azar y a la selección, si el estímulo persiste durante suficientes generaciones? A título personal, pienso que un modelo neolamarckista (que postula la herencia de caracteres adquiridos), bien desarrollado empíricamente, sería (y puede que algún día lo sea) el modelo ideal para explicar la adaptación de las especies, un modelo libre del azar y la competición y capaz de hacer frente a los problemas como el del ejemplo anterior
Sobre la herencia de caracteres adquiridos
En este apartado expondré una hipótesis sobre la herencia de caracteres adquiridos que sería capaz, al menos en parte, de explicar la adaptación de las especies al ambiente de forma lógica y plausible. Esta idea, pese a estar fundamentada en hechos empíricos, es solo eso: una idea. Existen tres estudios cruciales para la génesis de este modelo: La respuesta del genoma al ambiente, el descubrimiento de la variación estructural en los seres humanos y la hipótesis Soma-to-Germline de Ted Steele.
En primer lugar, es un hecho harto conocido el que los organismos son capaces de adaptarse al ambiente durante su vida, y que las respuestas involucradas en este hecho están dirigidas por el genoma. Ejemplos típicos son la reacción de la piel a la alta irradiación solar (que popularmente se conoce como “ponerse moreno”), la creación de masa muscular ante un ejercicio diario o el aumento de glóbulos rojos ante una situación prolongada de anoxia en montañas.
Por otra parte, hasta hace poco tiempo se pensaba que las variantes alélicas eran la única fuente de variabilidad dentro de una población, sin embargo, trabajos recientes e independientes ponen de manifiesto que gran parte (si no la mayoría) de las diferencias entre individuos no responden a diferencias alélicas de algunos genes sino al número variable de repeticiones que presentan de estos, principalmente relacionados con la respuesta al ambiente:
“Genome researchers now have a catch-all phrase for the vast array of rearrangements – including copy number polymorphisms, inversions deletions and duplications – that occurs normally in the human genome. They call it structural variation and have described at least 800 individual variants that, in total, account about the 3.5 % of the human genome. […] Eichler´s analysis this year showed that many of the genes found in structural variants negotiate our interactions with the environment”. (Erika Check, 2005).
Los últimos estudios se centrarán en buscar si existe variabilidad estructural entre las etnias humanas, y por tanto, si esta variabilidad está implicada en la adecuación de las poblaciones al ambiente.
Por último, Steele propuso en la década de los 70 un modelo para explicar un “extraño” suceso: la transmisión de información referente a regiones variables concretas de los anticuerpos del sistema inmune a la descendencia. Estas regiones mutan rápidamente ante un estado infeccioso para conseguir una variedad de anticuerpos muy afín a los agentes infecciosos (a sus antígenos en verdad), de manera que cuando esto se consigue se fabrican estos anticuerpos en grandes cantidades. Así, cada vez que un individuo se enfrenta a un nuevo agente ha de realizar cientos de pruebas hasta producir (si la llega a producir) la variante de anticuerpo específica y poder combatir la infección. Con cada infección, el organismo retiene una cierta memoria de manera que si se encuentra de nuevo con el mismo antígeno es capaz de producir anticuerpos de forma inmediata.
Steele comprobó que una parte de la descendencia de los individuos con los que trabajaba, nacían con capacidad de respuesta inmediata ante un antígeno que había sido inoculado a un parental con anterioridad. De hecho, la tolerancia a antígenos de otra cepa, que se puede conseguir al introducirlos durante los primeros días de vida del animal, se heredaba de forma incomprensible según la teoría de la barrera de Weissmann, según la cual la línea germinal y sómatica son totalmente independientes y, por lo tanto, es imposible la transmisión de caracteres adquiridos a la descendencia (figura adjunta 1, obtenida de “Lamarck´s signature”, Steele 1998).
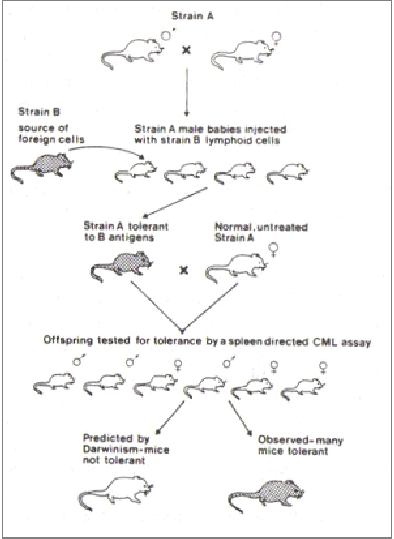
FIG.1: Experimento sobre la herencia de inmunidad adquirida. De “Lamarck´s Signature”, Steele 1998
Para explicar este hecho, Steele propuso su teoría, que llamó Soma-to germline loop, liderada por la actividad de partículas retrovíricas endógenas que serían capaces de transformar el RNA de los genes de los anticuerpos en manufacturación en los linfocitos B (línea sómatica) en DNA (mediante la enzima vírica transcriptasa inversa) y encapsular este material en una envoltura protéica, abandonar la célula y dirigirse a las gónadas (línea germinal). Finalmente las partículas víricas entrarían en las células sexuales pudiendo insertar la información que poseen en estas y confiriéndoles una prerrespuesta al antígeno (Figura adjunta 2). Este mecanismo, extraño pero coherente, es avalado por hechos como la alta presencia de retrovirus endógenos en los genomas (8% en nuestra especie), la posesión de transcriptasas inversas en virus y eucariotas (telomerasas), la creciente importancia de los elementos móviles en las hipótesis evolutivas y la similitud con los ciclos vitales de los retrovirus “libres”.
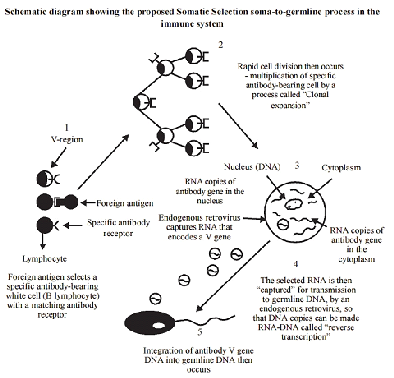
Figure 5. Schematic diagram showing the proposed ‘Somatic Selection’ soma-to germline process in the in the immune system proposed by Steele (1979) and modified from Steele et al. (1998), and to which we adhere for late metazoans with epigenetic development that possess a clearly differentiated germline. Such organisms developed a germline alter the primordia of major organ systems of the adult had become established (e.g. annelids). Our adherence to this Neo Lamarckian theory extends to the other extreme of the embryological iming that specifically includes the terminal differentiation of the germline, in organisms with preformistic development (e.g., mammals, bird, mice, and humans). Here the germline is terminally differentiated in earliest ontogeny, in which orientation of maternal-derived determinants deposited in the egg in present. 1. A diverse array of B lymphocytes exists before the entrance of a foreign antigen into the system. Each cell exposes on its surface membrane antibodies of one unique specificity. The variable (V)-region genes of the lymphocytes encode those parts of the antibody that form the antigen-combining site. A foreign antigen binds to those b cell which have compatible antibodies; therefore, these cells are ‘selected’ in a Darwinian manner, generally called clonal selection. 2. After antigen binding the B cell is activated and divides, producing progeny profusely that in turn divide, producing more identical progeny cells. A clone of identical cell expressing that antibody is produced (this is called clonal expansion). Individual cell of the clone can mutate their variable region genes (see Figures 2 and 6) that can themselves be selected by an antigen to produce another clone. 3. Within cell of the clone, RNA copies of antibody V genes are made in the nucleus. Mature mRNA is sent to the cytoplasm where it is translated into the appropriate sequence of amino acids making up the protein chains of the antibody. 4. RNA molecules encoding V genes (in the nucleus of the cytoplasm) may be engulfed by innocu-ous endogenous RNA retroviruses (produced by the cell) and complementary DNA copies of the RNA made by the viral enzyme, reverse transcriptase. These are the molecules that probably belong to the ancient RNA world and which were discovered by Temin in 1959 (see Temin, 1989) and by Temin and Baltimore in RNA tumor viruses in 1970. 5. According to Steele (1979) and to Steele et al. (1998) Neo-Lamarckian theory, which we adhere to for late Metazoan epigenetic organisms, and to the other extreme of the ontogenetic timing, for those with a preformistic mode of development, copies of the antibody V genes are transferred (alla Temin and Baltimore) by the virus with the inserted somatically mutated characteristic to sperm or egg nuclei. Thus, the germline DNA is enriched with more essential elements of the antibody repertoire. See a molecular unifying hypothesis (Bladen and Steele, 1998)
FIG. 2: Ver parte superior. Obtenida de: Genetics and Molecular Research 2 (1): 7-28 (2003) http://www.funpecrp.com.br/
Steele planteó no solo la posibilidad de herencia de caracteres en el caso del sistema inmune, sino que propuso varios ejemplos en los cuales, al igual que en el apartado anterior, la teoría neodarwinista no tenía sentido alguno: las callosidades de las patas de los avestruces, la modificación ósea de las articulaciones del tobillo en personas asiáticas y aborígenes australianos (adaptadas a su forma de sentarse) y por supuesto, la carencia de barrera de weissmann en los vegetales.
Ahora bien. ¿Cómo podría generarse la diferencia de caracteres? ¿Cómo podría reconocerse a los elementos del ADN implicados en estos? Recapitulando, tenemos la capacidad de adaptación del organismo al ambiente mediada por su genoma, la variabilidad estructural y la hipótesis Soma-to-germline loop. Veamos que surge si relacionamos conceptos y aplicamos lógica e imaginación. Propongamos una situación ideal: un organismo en condiciones ambientales normales produce de forma normal las proteínas necesarias, por medio de una actividad enzimática correcta y una regulación por parte de otras regiones del genoma. Ese mismo organismo, cuando se encuentra ante una situación de cierta irregularidad ambiental que persiste en el tiempo (entiéndase cualquier estímulo para el que no estaba originalmente preparado: alta irradiación solar, baja concentración de oxígeno, actividad muscular elevada…) reacciona adaptándose al ambiente variando la actividad de los genes o sus secuencias reguladoras, por ejemplo, aumentando drásticamente la producción de una proteína (como la melanina, que determina la pigmentación de la piel y su producción es mayor ante una alta irradiación solar, que es potencialmente mutágena). Así, cuando un estímulo incide sobre un organismo este responde adaptándose al ambiente. El estímulo persiste durante tiempo prolongado y el organismo sigue produciendo una alta cantidad de la proteína en cuestión. La alta actividad de transcripción podría ser detectada por un hipotético complejo, que llamaremos Sistema de Detección de Hiperactividad, capaz de unirse a regiones con una transcripción anormalmente elevada (por ejemplo siendo sensible a la concentración de elementos de transcripción o de GTP, a la aglomeración estructural de ribosomas, etc) y que portase un dominio transcriptasa inversa. De esta manera, cuando el SDH se une a una secuencia de ADN con una alta tasa de transcripción, la enzima transcritas inversa transcribiría las copias de ARN que está produciendo el gen en copias de ADN libres. Estas copias podrían ser encapsuladas, marcadas en el aparato de Golgi y dirigidas hacia las gónadas. Una vez alcanzada una célula sexual, la copia se insertaría en el genoma y como resultado la célula porta una copia extra del gen sobrexpresado en respuesta al ambiente, de manera que el individuo adulto resultante sería capaz de producir el doble de proteína sin entrar en estado de hiperactividad, sino que es el estado normal. De esta forma el descendiente heredaría la adaptación al ambiente del parental y, aunque desaparezca el estímulo, su fenotipo permanecería, a menos que la copia se inactivase por otros mecanismos. (Ver figura adjunta 3).
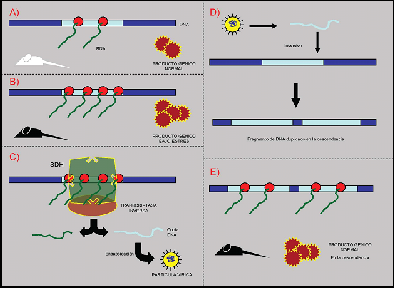
FIG. 3: Mecanismo propuesto para la transmisión de caracteres adquiridos. Izquierda: línea somática del parental. A) Producto normal del tipo parental. B) Producto en masa bajo estrés ambiental. C) Actividad del SDH-trascriptasa inversa, retrotrascripción y encapsidación de la copia del gen. Derecha superior: línea germinal del parental. D) Inserción de la copia produciendo una duplicación del gen. Derecha inferior: línea somática de la descendencia. E) Producto normal del tipo descendiente. (Daniel Heredia, 2005)
Conclusiones
La adaptación es un estado evidente de los seres vivos, pero la forma en que se produce solamente es explicada mediante la teoría neodarwinista bajo la selección y el éxito. Sin embargo, en poblaciones grandes, normales y fuera de un estado crítico anómalo que genere competencia (superpoblación, extinción de recursos, igualdad de nichos por introducción de especies alóctonas…), es difícil sostener que es este mecanismo el generador de la adaptación. Por lo tanto han de existir otros procesos distintos que den lugar a la adaptación de las especies, que además probablemente difieran dentro de los dominios y los reinos e incluso los phila. En este ensayo se presentó un modelo hipotético, aunque basado en estudios reales, para tratar de abarcar, hasta cierto punto, el proceso adaptativo de los animales.
Esta proposición es, por supuesto, divagadora y simple, puesto que es lógica la acción de intermediarios en la señalización, transcripción, inserción, etc.
No obstante, pese a ser “ideal”, esta idea es coherente con ciertos hechos como la existencia de transcriptasas inversas, regiones del genoma con alta producción y gran número de copias (NOR, genes de histonas), la adaptación de las poblaciones en ausencia de selección y la variación estructural de secuencias relacionadas con la respuesta al ambiente. La velocidad del proceso adaptativo sería variable, puesto que el llevar a cabo el proceso completo dependería de el correcto funcionamiento de todos los pasos. Igualmente, la “infección” no sería igual en todos los gametos, con lo que se crearía cierta variabilidad dentro de la descendencia.
Además, existen ejemplos bastante compatibles con este modelo, como el descubrimiento reciente de que en los pinzones de las Galápagos las diferencias morfológicas y adaptativas del pico están determinadas por la cantidad de proteína BMP4 presente y su momento de expresión durante el desarrollo. Este descubrimiento va de la mano con la curiosa capacidad de hibridar entre las distintas especies de pinzones de Darwin y dejar descendencia fértil (lo que lleva a la conclusión lógica, aunque aún no aceptada, de que pertenecen a la misma especie), lo cual no deja de ser una paradoja, pues fueron estos los pájaros sobre los que Darwin desarrolló su teoría del origen de las especies.
Las preguntas lógicas y cruciales que me surgen ante este modelo son: ¿Cómo se activa? ¿Como se regula? ¿Cómo se eliminan o silencian las copias? ¿Existen periodos de mayor sensibilidad?
Respecto a otros organismos, la ausencia de la barrera de Weissmann y la alta plasticidad que poseen plantas, hongos, protistas y bacterias, así como las características propias de cada uno (unicelularidad, corta vida, capacidad de hibridación, etc) no permite generalizar y su adaptación al medio debe regirse por leyes previsiblemente distintas entre ellas, del modelo aquí propuesto y de la teoría neodarwinista.
Las Fuentes de variación:
Mecanismos generadores biodiversidad
Tratar de explicar toda la biodiversidad bajo las premisas darwinistas es imposible siguiendo distintas evidencias científicas: paleontológicas (evidencias de saltacionismo), genéticas (gran complejidad genómica, imposibilidad de establecer la mutación puntual como fuente de cambio evolutivo) y demográficas (imposibilidad de establecer aislamiento reproductivo en situaciones simpátridas de forma gradual). Asumiendo este criterio y teniendo en cuenta la complejidad de la vida, la evolución ha de ser por lógica un proceso complejo y multifactorial, en el que se han debido ver involucrados distintos mecanismos de cambio rápido. A continuación se comentarán algunas posibles fuentes de cambio evolutivo capaces de dar lugar a la especiación y cambios en los organismos perpetuables en el tiempo, siendo independientes de la dinámica de las poblaciones y las premisas gradualistas.
Las mutaciones
Una mutación es por definición un cambio en una secuencia genética determinada. Estos procesos pueden ser espontáneos, producidos por errores en la replicación de manera aleatoria, o inducidos por agentes físicos y químicos mutágenos. Las mutaciones espontáneas son eventos raros en la naturaleza y es difícil obtener un gran número de mutantes para estudiar de poblaciones naturales. Las mutaciones pueden ser de distinta naturaleza, hablamos de mutaciones puntuales cuando es afectada una base y su complementaria (transiciones, transversiones, delecciones e inserciones). Debido a las propiedades del código genético, el que sea degenerado, las mutaciones pueden ser neutras en sus efectos porque un distinto triplete de bases puede codificar la entrada del mismo aminoácido. Aun en el supuesto de que el aminoácido cambie, las propiedades estructurales de la proteína final pueden conservarse y mantener su función, tal como puso de manifiesto Kimura en su teoría neutralista. Además, la increíble complejidad de las interacciones génicas, que ponen de manifiesto que el fenotipo de un carácter no depende de un gen sino de las distintas cantidades expresadas por múltiples genes que interactúan, establece que distintos genotipos producen un mismo fenotipo, complicando aún más la posibilidad de que un cambio puntual pueda afectar al individuo sustancialmente. Sin embargo, las mutaciones puntuales existen y en ocasiones producen efectos visibles: malformaciones, enfermedades o muerte de los individuos. Las mutaciones no neutras provocan la inactivación de un gen, afectando a la cascada de productos que dan lugar a un carácter, lo que da lugar a organismos mal desarrollados. Las “mutaciones beneficiosas” no han sido nunca registradas y son incoherentes con la propia naturaleza desorganizadora de las mutaciones y con la inmensa homología génica que presentan los genomas. Por todo ello, aunque han sido la base teórica de los cambios graduales, no haremos más hincapié en las mutaciones discretas como fuente de cambio.
Las mutaciones frecuentemente no son aisladas, sino que se concentran en lugares específicos del genoma, los hot-spots mutacionales (puntos calientes). En estos lugares la tasa de mutación es superior que en otras partes del genoma, siendo un fenómeno más corriente y prolífico. Existen mecanismos de tamponamiento mutacional, como la proteína HSP90, que inactivan las mutaciones durante generaciones, pero cuando estos sistemas se suprimen pueden expresarse en un individuo todas las mutaciones inactivas de forma brusca.
Pero además de las mutaciones discretas existen fenómenos macromutacionales vinculados a la dinámica del genoma, las que más nos interesan son las duplicaciones. Las duplicaciones son copias de un segmento de ADN que pueden ocurrir principalmente por sobrecruzamientos desiguales entre cromátidas homólogas. Según Ohno (1970): “Las duplicación del ADN es la mayor fuerza de la evolución”. Esta afirmación se basa en la posibilidad de destinar una de las copias del ADN para su función original mientras la otra puede “experimentar” nuevas funciones, generar el doble de producto génico y permite la especialización de enzimas multifuncionales en una sola actividad (perdiendo las otras).
El proceso de duplicación toma una dimensión distinta cuando la situamos en el contexto de los genes homeóticos o genes Hox. Estos genes son los organizadores del desarrollo embrionario, destinados a dirigir la morfología del individuo. Los genes Hox no generan las estructuras de forma directa, sino que las ordenan a lo largo del eje anteroposterior de los animales (en vegetales existe genes análogos llamados Madt). Estos genes determinan los diferentes segmentos del cuerpo regulando a otra serie de genes estructurales específicos en cascada (genes Downstream). Los genes homeóticos han sido bien conservados durante la evolución, de forma que es posible reconocer los mismos Hox en organismos muy distantes filogenéticamente. Pero es más, experimentos revelan que su funcionalidad se mantiene entre especies muy distintas: las mutaciones en el gen Deformed (Dfd) causan defectos fatales en la cabeza de D. melanogaster que puede ser “rescatada” por la acción de un gen equivalente humano, el Hox-4. Esta conclusión implica que tanto los genes Hox como los que regulan (downstream) han de estar bien conservados, y presumiblemente debe ser su regulación y las diferencias en lo genes estructurales las que originan estructuras distintas como las cabezas de mosca y humano. Cada gen Hox está ligado a un segmento concreto y existe cierta jerarquía genética entre estos. Los genes downstream parecen ser esencialmente los mismos para todos los Hox, lo que diferencia a unos de otros es la afinidad de las proteínas Hox reguladoras por los genes downstream.
Volviendo a las duplicaciones, los animales más simples con plan morfológico definido, los cnidarios, tienen un reducido número de genes Hox (en concreto, Hidra tiene 3) frente a los animales complejos bilaterales que tienen un número mayor. La duplicación parece un requisito para la aparición de los genes Hox sucesivos y su diferenciación. Cada Hox determina un segmento en el cuerpo de un animal, creando metámeros o somitos, la base de los animales metaméricos, como los anélidos, los artrópodos o los vertebrados. Los genes homeóticos y sus cascadas génicas representan una importante fuente para la variación de la forma, si bien un pequeño cambio en su regulación o expresión puede ocasionar grandes diferencias morfológicas.
Los reordenamientos cromosómicos
Las modificaciones del genoma no están sujetas al nivel génico, sino que pueden darse alteraciones de mayor índole, en los cromosomas o incluso el genoma completo. El proceso de duplicación también es el más interesante en este caso, aunque son importantes también las inversiones y delecciones. Existen trazas de duplicaciones de cromosomas concretos por divisiones meióticas incorrectas, así como el resto de los cambios estructurales de los cromosomas. Parece que este tipo de fenómenos han sido frecuentes durante la evolución y de hecho aparecen diferencias cromosómicas importantes entre especies tan próximas como los diferentes póngidos y humanos. Los accidentes cromosómicos producen una barrera reproductiva entre el organismo ancestral y los modificados, debido a los problemas que surgen en la recombinación meiótica (incapacidad de unirse al homólogo) y se reduce la fertilidad.
Por otro lado existen los fenómenos de duplicación completa de la dotación genética, por formación de gametos no reducidos. Este suceso parece tener una importancia constatada en la evolución de las plantas, como veremos cuando hablemos de la hibridación, y probablemente en animales. En el caso de estos últimos se conoce un ejemplo de poliploidación reciente (Beçak y Beçak, 1971) en Odontophrynus americanus, un anuro brasileño del que existen dos poblaciones, una diploide (2n) y otra teraploide (4n). Las diferencias entre las poblaciones son escasas, principalmente de tamaño, por lo que se las considera sibling species (especies hermanas, que presentan un claro aislamiento reproductivo, pero son morfológicamente casi iguales). Este hecho es interesante, porque pone de manifiesto una cuestión importante: la especiación y la adaptación o cambio morfológico no están irreversiblemente ligadas. Las sibling species son muy frecuentes entre los insectos. Las duplicaciones de genoma completo pueden haber sido de gran importancia en la evolución de los vertebrados, como en la radiación de los peces teleósteos (Venkatesh).
Los Elementos transponibles
Llamamos elementos móviles o transponibles (ETs) a aquellas secuencias genómicas capaces de trasladarse de un lugar a otro a través del cromosoma. Aunque fueron incluidos como ADN basura, actualmente se considera que pueden tener importancia para la movilización de exones y secuencias promotoras, la separación de genes duplicados y para el barajamiento general del genoma. La transposición es un fenómeno ubicuo, altamente responsable de la organización del genoma. Además, son una fuente de variación tanto las inserciones en secuencias codificantes o reguladoras, que afectan a la trascripción y los transcritos e inactivan genes, como las interacciones que se dan entre elementos homólogos (apareamiento ectotópico), que producen inversiones, delecciones, translocaciones o inversiones en los cromosómas (reordenamientos cromosómicos) y afectan o inactivan genes. La mayoría de las mutaciones morfológicas en Drosophila (80%) y el 15 % en mamíferos son producidos por la inserción de ETs. Una gran parte del ADN repetitivo disperso son ETs, de hecho, se estima que estos elementos representan el 50% del genoma del maíz, el 10-15% del de Drosophila y, posiblemente, al menos un 25% en los seres humanos.
Podemos distinguir tres tipos de elementos transponibles: transposones (ETsII), retrotransposones y retrovirus endógenos (en conjunto: retroelementos, ETsI). Además, existen otros elementos móviles poco corrientes y de difícil clasificación (ETsIII).
La Hibridación
La hibridación es literalmente el cruzamiento entre dos especies u otras categorías taxonómicas inferiores. Este es un fenómeno común en el mundo vegetal, donde las especies no garantizan el aislamiento reproductivo total y es una fuente muy importante para la especiación y evolución de las especies vegetales.
La hibridación suele ser asimétrica, una especie parental siempre aporta el gameto masculino y autoincompatible, y la otra el femenino y autocompatible. Las especies híbridas suelen prosperar con los cambios ambientales y en hábitats híbridos.
Los híbridos suelen presentar características intermedias de ambas especies, así como características nuevas emergentes debidas a la heterocigosis y las nuevas combinaciones génicas. Además los especimenes híbridos suelen crecer más vigorosos, lo que se denomina heterosis. Sin embargo, la fertilidad de estas plantas se ve reducida con la hibridación, con lo que están limitados aunque puedan generar nuevas especies. Para ello han de recuperar su potencialidad, bien manteniendo el número de cromosomas original (especiación híbrida homoploide e hibridación introgresiva) o bien por medio de poliploidía (especiación alopoliploide). No obstante, los híbridos estériles pueden reproducirse asexualmente por apomixis.
La especiación híbrida homoploide requiere de recombinación por autofecundación mediante cruzamiento entre sí o con las especies progenitoras. Este proceso está asociado a rápidos cambios cromosómicos por mutaciones posteriores, lo que establece un aislamiento reproductivo. Experimentos con girasoles híbridos (Helianthus anomalus) muestran un aumento de la fertilidad de menos del 10% al normal en cinco generaciones.
La hibridación introgresiva es un proceso producido por el retrocruzamiento continuo o muy frecuente, lo que provoca la permanencia de solo unos pocos caracteres de un parental respecto del otro. Esto genera poblaciones de un parental con nuevos caracteres procedentes de otra la otra especie, sin establecerse aislamiento reproductivo, sino más bien un proceso indirecto de transferencia de genes entre especies por medio de la descendencia híbrida.
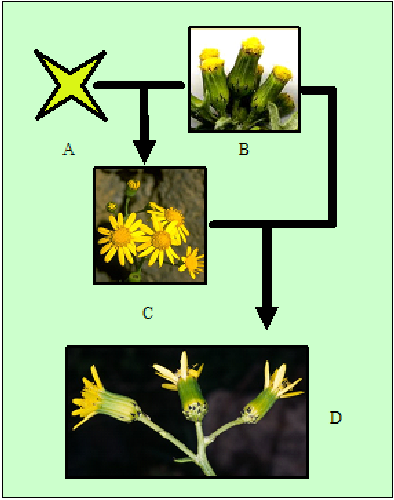
FIG. 1: Hibridación introgresiva en Senecio sp. La especie híbrida S. squalidus (C) presenta flores liguladas de gran tamaño, mientras que la especie ancestral S. v. vulgaris (B) no tiene flores liguladas. Del cruzamiento introgresivo de estas especies aparece la variedad S. v. hibernicus (D) de la especie ancestral, que tiene flores liguladas cortas. A) La otra especie parental de S. squalidus.
La alopoliploidia consiste en la recuperación de la fertilidad (reducida por mecanismos de aislamiento postcigótico) por medio de poliploidación. Tras la formación del híbrido, la formación de bivalentes en el proceso meiótico de la planta para la reproducción se ve inhibida en mayor o menor medida por la falta de homología entre las cromátidas. Si se produce la fusión de gametos no reducidos (con dotación 2n en vez de n) se formará un híbrido poliploide, en el que cada cromosoma tiene un homólogo con el que puede recombinar, y se recupera la fertilidad, Triticum aestum y Polipodium son ejemplos de plantas con alopoliploidia estudiadas. La alopoliploidia concede aislamiento a los híbridos y los retrocruzamientos producen triploides con problemas de fertilidad. La frecuencia de aparición de especies híbridas aumenta con la latitud (floras escandinavas presentan 56%-72% especies poliploides, floras mediterráneas 23%-34%), hecho que puede estar relacionado con el cambio climático del cuaternario (fin de la glaciación Würm) y la aparición de nuevos ambientes terrestres.
La hibridación en animales es un proceso mucho más raro y conlleva la esterilidad de los individuos, debido a que no posemos la plasticidad de los vegetales ni su versatilidad. No obstante, en algunas especies la hibridación es un fenómeno posible: el pez Gila seminuda es de origen híbrido entre las especies G. robusta y G. elegans (Dowling y DeMarais, 1993); el género de insectos palo Bacillus contiene varias especies de origen híbrido también. Los animales híbridos suelen ser fruto de alopoliploidía y adoptan sistemas de reproducción clonal (como la partenogénesis).
La Simbiosis
La simbiosis es un proceso de integración o cooperación entre dos especies. Este hecho a priori parece tener poco que ver con la biología evolutiva, pero de hecho, la simbiosis es un fenómeno de gran importancia en la historia y configuración de la vida en la tierra.
Así pues, el mayor avance evolutivo de todos los tiempos (junto con la aparición de los distintos bauplanes durante la explosión cámbrica) es producto de la simbiosis: la célula eucariota. Este hecho fue propuesto por Lynn Margulis en 1967 en el Journal of Theorical Biology, tras el rechazo previo de varias revistas. La teoría de la endosimbiosis serial de Margulis dicta que la primera célula eucariota surgió como producto entre la fusión inicial de dos bacterias al que se le sumaría la presencia endosimbionte de otros tipos bacterianos. El primer paso fue la unión de una bacteria del dominio archaea con una espiroqueta (eubacteria) seguida de la fusión de sus genomas, que quedarían empaquetados en un núcleo. La unión de la espiroqueta aportó, a parte de su información genética, componentes del citoesqueleto celular e incluso un flagelo derivado del cuerpo celular adherido a la archaea. Tras este primer paso, a este cuerpo central de la célula se unirían las bacterias precursoras de cloroplastos y mitocondrias, que establecerían una relación simbionte que perdura hasta nuestros días, permaneciendo en el citoplasma celular e insertando la mayoría de sus genes en el núcleo del hospedador.
Además, multitud de formas protistas parecen haber surgido por medio de endosimbiosis posteriores, por lo que este es un mecanismo que ha aparecido varias veces en la evolución (figura 2).
Existen multitud de simbiosis de gran importancia en el mundo animal, que confieren beneficios adaptativos a las especies, por lo que la importancia de la simbiosis parece digna de tener en cuenta.
En el mundo vegetal existe un importante grupo derivado la simbiosis entre organismos distantes: los líquenes. Estos son el resultado de la unión entre hongos y algas unicelulares de distintas especies. Los líquenes son un grupo prolífico y muy versátil que crece en superficies rocosas y troncos vegetales, siendo de gran importancia ecológica. Aunque no existe una fusión física de los genomas, estos actúan en conjunto pero independientes, dando lugar a un organismo autónomo.
Por último, queda mencionar la llamada Simbiogénesis, es decir, la especiación mediada por la adquisición de simbiontes. Aislamiento reproductivo de una población se produce por la presencia de un simbionte (en principio) por parte de unos organismos pero ausente en otros. Este es el caso de Wolbachia, una bacteria que puede estar presente en distintas especies de insectos provocando el aislamiento reproductivo de sus hospedadores por incompatibilidad citoplasmática.

FIG 2: Simbiosis establecidas en la diversificación de los protistas.
Evolución, Especiación y Adaptación:
Visión general personal
Tradicionalmente se consideran a los fenómenos de adaptación, especiación y evolución como un proceso continúo derivado de la actuación de la selección natural sobre las poblaciones, las mutaciones puntuales que aparecen en su seno y la reproducción diferencial de los más adecuados. Sin embargo, es un error inaceptable considerar bajo una lupa reduccionista la diversidad de la vida, admitiendo que todo se debe a la extrapolación de un único y omnipotente proceso de gran simplicidad.
Además, los análisis procedentes de distintas disciplinas establecen diferencias entre la adaptación, la especiación y la evolución (comúnmente llamada “macroevolución” debido al caos terminológico en que nos situamos).
He aquí mi visión personal de tres aspectos fundamentales, relacionados pero no concatenados.
Adaptación
La adaptación es el proceso por el cual los organismos y las especies responden al ambiente y se adecuan a este, permitiendo la supervivencia de los/las mismas frente a cambios en su ambiente.
La adaptación implica pequeñas variaciones en la morfología y fisiología de los organismos que pueden ser o no heredables. Estas variaciones pueden apreciarse en mayor o menor medida, pero en ningún caso pueden dar lugar a la aparición de especies nuevas. Este ajuste de las poblaciones es, sin embargo, fundamental para la vida y puede explicar algunas características claramente ligadas al entorno de los organismos.
La adaptación no heredable se debe a la capacidad de respuesta al ambiente que presentan los seres vivos, que permite una relativa, pero fundamental, plasticidad ante cambios ambientales y puede dar lugar a ecotipos (conjunto de organismos de una especie con características particulares, no heredables genéticamente, en respuesta a un ambiente concreto).
Por otra parte, la adaptación con resultados genéticamente heredables ha sido y es el principal fenómeno estudiado en relación a la adaptación de las especies en términos “evolutivos”. Desde el punto de vista neodarwinista, la adaptación es el resultado de la actuación de la selección natural, es decir, la supervivencia de los más adecuados en cada ambiente y momento. Este supuesto, no ocurre de forma natural en las poblaciones por medio de la competencia, sino que en todo lugar sería el resultado de un efecto de cuello de botella, en el que la aparición de un efecto nocivo limitante produce un sesgo en la población. Sin embargo, aunque podría decirse que la población se ha “adaptado” a la nueva condición, en verdad no ha aparecido ninguna novedad para hacer frente al problema, sino que han sobrevivido caracteres preexistentes que formaban parte de la variabilidad de la población. De igual modo, otras modalidades de deriva genética (efectos insulares y fundadores) pueden explicar ciertos rasgos heredables de una población, aunque en realidad no se pueda considerar una verdadera adaptación.
Sin embargo, existen numerosos casos concretos que no pueden ser explicados mediante procesos de reducción de la variabilidad existente en pos a la supervivencia de la especie, sino que deben haber requerido la implicación de mecanismos concretos de adaptación heredable por respuesta al ambiente. De esta forma, las poblaciones se adaptarían como consecuencia de la adaptación al unísono de los organismos que la componen en respuesta al ambiente. El problema fundamental consiste en encontrar el mecanismo viable capaz de provocar que dichos cambios sean heredables, como son la mayoría de las características de tipo adaptativo. Así pues, podrían ser candidatos a este puesto fenómenos epigenéticos, de supresión génica y duplicaciones selectivas de genes estructurales y reguladores.
La aparición de nuevos caracteres por mutaciones compatibles con la vida (pigmentación principalmente) podría provocar, en casos concretos, un efecto inverso al de adaptación, en el que los organismos anómalos deberían encontrar un ambiente propicio (para la cripsis) o morir.
La adaptación, en resumen, permite la supervivencia de las especies, pero no su formación. Los cambios que se dan lugar en los organismos y las poblaciones son reversibles, no heredables, o preexistentes en la variabilidad de la especie o producen pequeños cambios genéticos incapaces de acumularse o generar, de forma general, una incompatibilidad reproductiva real.
Especiación
La especiación es, literalmente, la formación de nuevas especies a partir de otras preexistentes.
Un problema fundamental consiste en definir el concepto de especie. Así pues, en los organismos con reproducción sexual, esta categoría implica siempre, además de cierto parecido morfológico, un aislamiento reproductivo total (animales) o relativamente alto (plantas) con respecto a todos los individuos no pertenecientes a su especie. En organismos procariotas, se establece como especie a aquellos conjuntos de bacterias con una homología genética superior al 97%, aunque por supuesto este es un criterio artificial, al igual que la separación de bacterias en especies. Por esto último, al hablar de especiación de aquí en adelante sólo se considerará a los organismos con reproducción sexual.
La visión neodarwinista afirma que las mutaciones puntuales, surgidas al azar y favorecidas por la selección natural, se acumulan en las poblaciones generando diferencias que, con el tiempo, acaban por establecer incompatibilidad genética para la reproducción con otros grupos poblacionales de su misma especie. Sin embargo, la acumulación de nuevos caracteres como vía de formación de las especies es más que cuestionable, debido a que mientras existe compatibilidad reproductiva entre los individuos, estos caracteres tienden a disgregarse o difundirse en la población, no creando aislamiento alguno y, en el mejor de los casos, aumentan la variabilidad de la especie creando polimorfismos. Así pues, la especiación simpátrida, por lógica la de mayor frecuencia, carece de credibilidad bajo las premisas neodarwinistas.
La especiación requiere de aislamiento reproductivo, y este a de darse de forma prioritaria por una barrera de aparición brusca. Los reordenamientos cromosómicos y del genoma completo provocan un aislamiento reproductivo postcigótico inmediato, y como resultado de un cruce con el genotipo ancestral o emparentado surgen híbridos estériles o abortivos. Los análisis cariológicos demuestran que existen reordenamientos cromosómicos están ligados especies emparentadas, como en nuestro caso, Homo sapiens, donde existe una homología genética de más del 98% con los chimpancés (escasas diferencias genéticas) pero se si observan claros reordenamientos en varios cromosomas, al igual que con otros grandes simios:
“Except for differences in non genetic heterochromatin, chromosomes 6, 13, 19, 21, 22, and X have identical banding patterns in all four species. Chromosomes 3, 11, 14, 15, 18, 20, and Y look the same in three of the four species (those three being gorilla, chimps, and humans), and chromosomes 1, 2p, 2q, 5, 7 - 10, 12, and 16 are alike in two species. Chromosomes 4 and 17 are different among all 4 species. Most of the chromosomal differences among the four species involve inversions - localities on the chromosome that have been inverted, or swapped end for end. This is a relatively common occurrence among many species, and has been documented in humans”.
Además, las regiones implicadas en reordenamientos parecen estar asociadas con duplicaciones a gran escala.
De igual manera, la poliploidización genera aislamiento postcigótico inmediato. Este fenómeno es común en plantas e insectos, aunque también se conocen casos en vertebrados (anuro Odontophrynus americanus) que dan lugar a “poblaciones con aislamiento reproductivo simpátrido”. Analizando los genomas de los teleósteos se ha concluido que las duplicaciones completas de genoma pudieron ser fundamentales en la radiación de estos peces.
Por otra parte, pueden formarse barreras reproductivas precigóticas que impiden que se produzca la fecundación. En los erizos de mar, existe una gran especificidad entre los receptores de huevos y espermatozoides, así como de las moléculas quimiotácticas que permiten el encuentro de los gametos. Las especies emparentadas mantienen características similares, aunque la hibridación es evitada por estos mecanismos. La aparición de una nueva variedad de señalización implicaría un aislamiento reproductivo.
En todo caso, estos mecanismos carecen de utilidad bajo las premisas neodarwinistas, donde las variaciones viables surgen en un sólo individuo. La aparición de cualquier mecanismo de aislamiento inmediato en un único individuo traería como consecuencia la incapacidad para reproducirse con ningún otro espécimen y, por supuesto, no se formaría especie alguna. Por ello, la especiación es un proceso que requiere una reorganización análoga en varios individuos, ya sea por que se de en individuos clónicos, inducida por influencias ambientales en una generación o por existir una predisposición estructural de que ocurra ese cambio concreto.
La especiación es un fenómeno que incrementa la diversidad de especies de un taxa, pero no necesariamente la disparidad, puesto que no siempre conlleva grandes cambios estructurales en la biología de los organismos (evolución). Tras la formación de una especie, a parte de los cambios derivados de la reorganización cromosómica, pueden acumularse características (ahora incapaces de mezclarse con la población original), procedentes de la adaptación y la deriva genética, que conforman diferencias más o menos leves (aunque apreciables) con respecto a otras especies emparentadas. Hablando coloquialmente, la especiación per se produce “variaciones sobre un mismo tema”, como las que se observan entre las especies de saltamontes, antílopes o pinos. La aparición de nuevos patrones biológicos (nuevos “temas”), aunque también lleve implícito el aislamiento reproductivo, es lo que podemos considerar, en sentido estricto, evolución.
Evolución
La evolución es el cambio de un estado a otro, la aparición de planes biológicos completamente nuevos, claramente distintos de los organismos precursores. Evolución implica un cambio representativo en la organización general de un patrón y que origina nuevos taxones reconocibles.
La evolución se establece como un proceso gradual de trasgresión de las especies, dando lugar a formas nuevas por medio del continuo proceso adaptación-especiación-evolución. Sin embargo, esta serie no parece tener validez real. Por otra parte, el registro fósil no muestra infinitas formas intermedias, como sería de esperar, sino especies definidas que perduran durante unos millones de años para desaparecer y ser suplantadas por otras de forma abrupta (el Equilibrio puntuado de Elredge y Gould).
Además, los grandes cambios evolutivos parecen estar relacionados con eventos de extinción de gran magnitud, achacados principalmente a fenómenos de gran inestabilidad ambiental. Tras estos fenómenos es corriente que se observen en el registro fósil “radiaciones adaptativas”, es decir, la aparición rápida de una gran cantidad de taxones nuevos. La explicación “ortodoxa” a este fenómeno es contradictoria con las propias premisas neodarwinistas: tras una extinción masiva, muchos nichos ecológicos quedarían “vacíos” y la ausencia de competencia favorecería la rápida, pero gradual, adaptación (?) de las especies dando como resultado radiaciones adaptativas. ¿Cómo es posible que si la competencia es el brazo ejecutor de la selección natural, motor de la evolución, su ausencia de lugar a la rápida aparición de gran cantidad de formas nuevas?
La evolución ha de ser, al igual que la especiación, un fenómeno rápido y que afecte por igual a varios individuos. Sin embargo, además deben aparecer grandes cambios biológicos en los individuos, resultantes de un mayor reordenamiento del genoma. Las reorganizaciones se deben no solo a la redistribución de cromosomas, sino a otros fenómenos como la asimilación de información foránea (simbiosis y transferencia horizontal genética) y, principalmentalmente, a la actividad de los elementos transponibles y otros elementos repetidos no codificantes de proteínas.
La importancia de los elementos transponibles ha sido ignorada hasta la década de los 90 debido a su prejuiciosa calificación como ADN basura, debido a que no codifican proteínas. Posteriormente se les tachó retorcidamente de “ADN egoísta”, por su insólita capacidad de moverse y copiarse a lo largo de los cromosomas (lo que se quiso interpretar como un “esfuerzo” por perpetuarse como polizones de lo genomas en que residen). Sin embargo ahora se empieza a reconocer la gran importancia de estas secuencias en los genomas eucariotas y en su evolución. Las secuencias repetidas no codificantes de proteínas, constituidas o derivadas de elementos transponibles en gran medida y abundantes en los genomas eucariotas, parecen ser fundamentales en muchos aspectos: constituyen las regiones pericentroméricas de los cromosomas, permiten la movilización de exones y secuencias promotoras, son fundamentales para la estructura de los cromosomas, contienen secuencias no codificantes de proteínas pero sí de RNAs que intervienen en la regulación y supresión génica; en nuestra especie, docenas de genes humanos parecen haber derivado de elementos transponibles y unos cientos de genes usan las terminaciones transcripcionales de retrotransposones del tipo LTR, otros genes usan los elementos reguladores derivados de elementos repetidos…
Pero además, es un hecho constatado la influencia del estrés ambiental en estas secuencias, produciendo su activación y provocando grandes duplicaciones, pero también inversiones, deleciones, translocaciones e inversiones cromosómicas debidas a la interacción entre elementos homólogos (apareamiento ectotópico). Parece ser que existen, además, sitios preferenciales de inserción, estableciendo cierta variabilidad predeterminada que puede influir en la probabilidad de que un mismo suceso ocurra en varios individuos por separado.
Por otra parte existen evidencias de la gran importancia que ha tenido la integración de información procedente de otros sistemas biológicos por medio de simbiosis celular, como la aparición de las células eucariotas y de gran cantidad de protistas posteriores, y por transferencia horizontal por medio de vectores víricos. Con su reactivación, un retrovirus puede llevarse consigo parte del ADN del lugar de inserción y propagarla por sus nuevos huéspedes, pudiendo traspasar la barrera de la especie.
En referencia a este último caso, el análisis inicial del genoma humano (2001) reveló que cientos de genes humanos parecen ser el resultado de transferencia horizontal desde bacterias en algún punto del linaje vertebrado. Muchas de estas nuevas adquisiciones han establecido papeles fundamentales en nuestro metabolismo (hidrolasas, deshidrogenasas, MAO). (Figura 1). Observaciones similares en el genoma de la lombriz indican una probable transferencia horizontal linaje-especifica.
Recapitulando, los grandes eventos extincionales y el gran estrés ambiental al que están ligados podrían ser los desencadenantes de los fenómenos evolutivos por medio de la activación de elementos móviles dentro de los genomas y fuera, en su forma libre de retrovirus. Así pues, se producirían grandes reordenamientos en los genomas que afectarían a su estructura interna y a su regulación general. El resultado final sería en muchos casos abortivo, al fin de al cabo se trata de una extinción, y no quedaría constancia fósil de estos intentos fallidos. No obstante, los organismos viables presentarían características notablemente diferentes a las de sus progenitores, y puesto que existe cierta variabilidad predeterminada en las inserciones de elementos móviles, es de esperar que más individuos presenten sus mismas características y la nueva especie pueda perpetuarse. Finalmente, la viabilidad de las nuevas formas dependería de su compatibilidad con la vida y la capacidad de integrarse en la compleja red de relaciones ecológicas de su ecosistema.
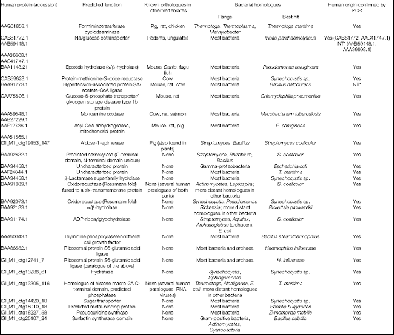
FIG. 1: Listado de genes bacterianos en humanos transmitidos por transferencia horizontal. Análisis inicial del genoma humano (2001)
|
Human protein (accession) |
Predicted funtion |
Known orthologues in other vertebrates |
Bacterias |
homologues |
Human origen confirmed by PCR |
|
|
|
|
Range |
Best hit |
|
|
|
|
|
|
|
|
|
AAG0853.1 |
Formiminotransferase |
Pig, rat, chiken |
Thermotoga, Thermoplasma, Methylobacter |
Thermotoga maritima |
Yes |
|
CAB81772.11 |
Na/glucose cotransporter |
Rodent, ungulates |
Most bacteria |
Vibrio parahaemolyticus |
Yes (CAB81772, AAC41747.1) NT* (AAB59448.1, AAA36608.1) |
|
AAA36608.1 |
Epoxide hydrolase (α/β-hydrolase) |
Mouse, Danio, fugu, fish |
Most bacteria |
Pseudomonas aeruginosa |
Yes |
|
CAB59628.1 |
Protein-mthionine -S-oxide reductase |
Cow |
Most bacteria |
Synechocystis sp. |
Yes |
|
BBA91273.1 |
|
Mouse, rat, cow |
Most bacteria |
Bacillus halodurans |
NT* |
|
CAA75608.1 |
Glucose-6-phosphate transporter/ glycogen storage disease type 1b protein |
Mouse, rat |
Most bacteria |
Chlamydophila pneumoniae |
Yes |
|
AAA59548.1 |
Monoamide oxidase |
Cow, rat, salmon |
Most bacteria |
Mycobaterium tuberculosis |
Yes |
|
AAF12736.1 |
Acyl-CoA dehydrogenase |
Mouse, rat, pig |
Most bacteria |
P. aeruginosa |
Yes |
|
AAA51565.1 |
Adose-1-epimerase |
pig (also found in plants) |
Streptomyces, Bacilus |
Streptomyces coelicolor |
Yes |
|
BBA92632.1 |
Predicted carboxylase (C-terminal domain, N-terminal domain unique |
None |
Streptomyces, Rhizobium, Bacilus |
S. coelicolor |
Yes |
|
BAA34458.1 |
Uncharacterized protein |
None |
Gamma-proteobacteria |
Escherichia coli |
Yes |
|
AAF24044.1 |
Uncharacterized protein |
None |
Most bacteria |
T. maritima |
Yes |
|
BAA34458.1 |
β-Lactamase superfamily hydrolase |
None |
Most bacteria |
Synochocystis sp. |
Yes |
|
BAA91839.1 |
Oxidoreductase (Rossmann fold) fused to a six-transmembrane protein |
None (several human paralogues of both parts) |
Actinomycetes, leptospira: more distant homologues in other bacteria |
S. coelicolor |
Yes |
|
BAA92073.1 |
Oxidoreductase (Rossmann fold) |
None |
Synochocystis, Pseudomonas |
Synechocystis sp. |
Yes |
|
BAA92133.1 |
α/β-hydrolase |
None |
Rickettsia; more distant |
|
Yes |
|
BAA91174.1 |
ADP-ribosylglycohdrolase |
None |
Streptomyces, Aquifex Archaeoglobus (archaeon) |
S. coelicolor |
Yes |
|
AAA60043.1 |
Thymidine phosporylase/endothelial cell growth factor |
None |
Most bacteria |
Bacillus stearothermophilus |
Yes |
|
BAA86552.1 |
Ribosomal protein S6-glutamic acid ligase |
None |
Most bacteria and archaea |
Haemophilus influenzae |
Yes |
|
GI_M1_ctg12741_7 |
Ribosomal protein S6-glutamic acid ligase (paralogue of the above) |
None |
Most bacteria and archaea |
H. influenzae |
Yes |
|
GI_M1_ctg13238_61 |
Hydratase |
None |
Synechocystis, Sphingomonas |
Synechocystis sp. |
Yes |
|
GI_M1_ctg13305_116 |
Homologue of histone macro-2A C-terminal domain, predicted phosphatase |
None (several human paralogues RNA viruses) |
Thermotoga, Alcaligenes, E. coli, more distant homologues in other bacteria |
T. maritima |
Yes |
|
GI_M1_ctg14420_10 |
Sugar transporter |
None |
Mos bacteria |
Synechoocystis sp. |
Yes |
|
GI_M1_ctg16010_18 |
Predicted metal-binding protein |
None |
Mos bacteria |
Borrelia burgdorferi |
Yes |
|
GI_M1_ctg16227_58 |
Pseudouridine synthase |
None |
Mos bacteria |
Zymomonas mobilis |
Yes |
|
GI_M1_ctg25107_24 |
Surfactin synthetase domain |
None |
Gram-positive bacteria, Actinomycetes, Cyanobacteria |
bacillus subtilis |
Yes |